|
STAGE
|
MITOSIS
|
MEIOSIS
|
|
Prophase
|
– chromosomes not visible
– Homologous chromosome remain separate
– No chiasmata formation
|
– chromosomes visible
– Homologous chromosomes pair up
– chiasmata formation take place
|
|
Metaphase
|
– chromaitd pairs line up on the equator of the spindle centromores line up on the equator
|
– this occur in metaphase II but no metaphase I
– centromere line equidislent aove and below the equater of the spindle
|
|
Anaphase
|
-chromatids separate number of chromosome present as parent cell
– Separated chromatids identical
|
– chromosomes separate in Anaphase I, chromatids separate in Anaphase II
– The separated chromosomes and chromatids may not be identical
– Half the number of chromosomes is present in daughter cell
|
|
Telophase
|
– Both homologous chromosomes are in each daughter cell
|
– only one of each pair of homologous chromosome is in each daughter cell
|
|
Occurance
|
– occurs in the formation of somatu cells
|
– Occurs in the formation of gametes and spores
|
STAGES OF SEXUAL REPRODUCTION
Sexual reproduction involves the following stages:-
1) gametogenesis
2) Copulation
3) Fertilization
4) Cleavage
5) Implantation
6) Pregnancy
7) Parturition(birth)
8) Parental case.
GAMETOGENESIS.
Definition:
Gametogenesis is the general process of gametes formation in both male and female reproducing sexually.
Meiosis is the process by which gametes are formed can also be called gametogenesis literally ‘creation of gametes’.
The type of meiosis in male organism forms a spermatogonium to a primary spermatocyte a secondary spermatocyte a spermatid and finally a spermatozoid is spermatogenesis.
Definition:
Oogenesis is the process of meiosis in female organism from oogonium to a primary oocyte, a secondary oocyte and then an ovum (egg cell).
The primordial germ cells once they have been populated the gonalds proliferate into sperm (in testes) or ova (in the ovary).
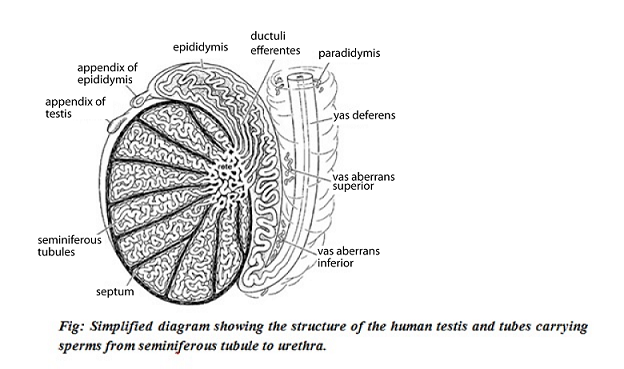
SPERMATOGENESIS
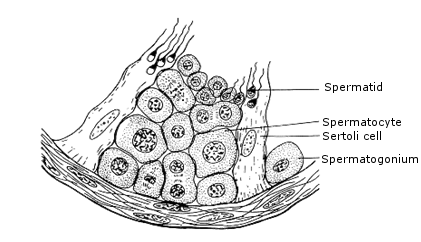
In male testis there are tiny tubules (seminiferous tubules) containing diploid cells called spermatogonia that develop into mature spermatozoa (spermatozoa are the mature male gametes in many sexually reproducing organisms).
In spermatogenesis i.e. a process during which spermatogonia (sperm cells) multiply giving rise to other spermatogonia restoring their population and to other which mature to spermatocyte.
Around the periphery of the seminiferous tubules are located specialized cells known as spermatogonia.
Spermatogonia are diploid cells set aside early in embryonic development. They may divide by mitosis to generate more spermatogonia or by meiosis to produce spermatids each of which will differentiate into a mature sperm cell.
Spermatogonia destined to undergo meiosis first differentiate into primary spermatocytes which undergo two successive meiosis divisions.
After meiosis I the produced cells are called secondary spermatocyte which each in turn undergoes the secondary division become spermatids each containing a unique set of 23 single chromosomes that ultimately mature into four sperm cells (spermatozoa).
The seminiferous tubular contain two types of cell;
Germ cells; these undergo the two division of meiosis to form the spermatozoa
Sertoli Cells: Acts as nerve cells ensuring that the germ cells have adequate nourishment.
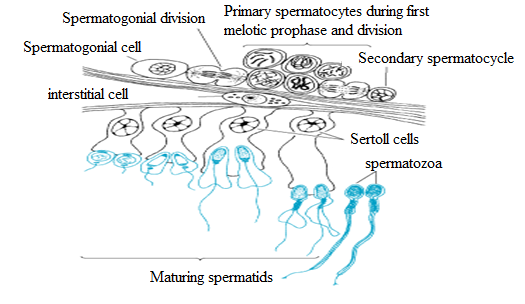
Fig: The stages of spermartozoa formation.
Spermatids undergo transformation in order to become spermatozoa
Fig: Diagram showing the structure of part of the wall of seminiferous tubule.
SPERMATOGENESIS
Occurs in seminiferous tubules.
Stored in epididymis.
Process:
Diploid spermatogonia divide by mitosis from germinal epithelium (germinal epithelial cells).
Some of them grow to produce diploid primary spermatocytes.
Diploid spermatocytes undergo first meiotic division to form two haploid secondary spermatocytes.
Haploid secondary spermatocytes undergo second meiotic division to form haploid spermatids.
These grow in shape and become spermatozoan.
The sertoli cells provide nutrition and protection against body immune system.
From the figure
The interior of the testis, site of spermatogenesis within the seminiferous tubules of the testis cells called spermatogonia develop into sperm, passing through spermatocyte and spermatid stages. Each
sperm passes as a long tail coupled to a head which contain a haploid nucleus.
MECHANISM OF SPERMATOGENESIS
The process of spermatogenesis is divided into the following phases (as shown below):-
1. Multiplication phase.
2. Growth phase.
3. Maturation phase.
4. Metamorphosis.
1. MULTIPLICATION PHASE
Also known as spermatocytogenesis.
Here the sperm mother cells present in the germinal epithelium of the seminiferous tubules divide repeatedly by mitosis to form a large number of diploid rounded sperm mother cells called spermatogonia.
Some of these sex cells move towards the lumen of seminiferous tubules and enter growth phase. These cells are called primary spermatocytes. The primary spermatocytes are diploid and contain (44 + XY)
chromosomes.
Some of these cells produced by the division of spematogonia remain in the original condition and continues to divide giving rise to primary spermatocytes such cells are known as stem cells.
GROWTH PHASE
During this phase, spermatocyte as well as its nucleus enlarges in size. It gets ready to undergo meiotic division.
3. MATURATION PHASE
Each diploid primary spermatocyte undergoes meiosis I which is a reduction division.
Two daughter cells are formed with ‘n’ number of chromosomes. The daughter cells are called secondary spermatocytes are haploid and much smaller comparatively containing (22 + X) or (22 + Y)
chromosomes.
The secondary spermatocyte undergoes the second meiotic division (equational). This results in the formation of four daughter cells known as spermatids
4. METAMORPHOSIS.
The spermatids formed as a result of maturation division in a typical animal cell with all the cell organelles present in it. In this form it cannot function as a male gamete. So many changes take place to change
the non – motile spermatid into motile spermatozoa.
The main aim of the changes is to increase the motility of the sperm. These changes are:-
Nucleus shrinks by losing water and DNA becomes closely packed.
An acrosome is formed from the Golgi complex.
An axial filament of the tail of the spermatozoa is formed from the distal centriole of the spermatid.
Mitochondrial ring is formed from the mitochondria around the distal centrioles and is called.
Much of the cytoplasm of the spermatid is lost and the remaining cytoplasm forms a sheath around the mitochondrial spiral. This is known as manchette.
During the process of differentiation, the developing sperms have their head embedded in the sertoli cells which are thought to provide nutrition for the developing sperms because their cytoplasm contains large stores of glycogen which diminish as spermatid mature.
NB: There is no direct evidence for this nutritional function of the sertoli cells, but some sperms of male sterility are associated with the failure to product normal sertoli cells]
Cellular events in human spermatogenesis
Sertoli cells support the developing gametes in the following ways:-
Maintain the environment necessary for development and maturation via the blood test is barriers.
Secretes substances initiating meiosis.
Secretes supporting testicular fluid.
Secrete the androgen – binding protein which concentrates high quantities of testosterone in close proximity to the developing gametes.
Testosterone is produced by intestinal cells (leydig cells) which reside adjacent to the seminiferous tubules.
Secrete hormones affecting pituitary gland control of spermatogenesis, particularly the polypeptide hormone, inhibin.
Phagocytise residual left over from spermiogenesis.
Release anti – mullerian hormone (AMH) which prevents formation of the mullerian duct/oviduct.
NB: Seminiferous epithelium in sensitive to elevated temperature in humans and will be adversely affected by temperature as high as normal body temperature.
Consequently, the testes are located outside the body in a sack of skin called the scrotum. The optimal temperature is maintained at 20C (man) -80 C below body temperature.
This is achieved by regulation of blood flow and positioning towards and away from the heat of the body by the cremasteric muscle and dartos smooth muscles in the scrotum.
Dietary deficiency (such as vitamins B, E and A), anabolic steroids, metals (calcium and lead) X – ray exposure, dioxin, alcohol and infectious diseases will also adversely affect the rate of spermatogenesis.
The hormonal control of spermatogenesis varies among species. In humans, the mechanisms are not known completely, however, it is known that initiation of the spermatogenesis occurs at puberty due to the interaction of the hypothalamus pituitary gland and leydig cells.
The hormones that are closely related to spermatogenesis are the lutenizing hormone, the follicle stimulating hormone (FSH) and testosterone (T).
LH controls spermatogenesis via the secretion of testosterone by leydig cells (3, 4, 5). Testosterone mainly acts onto sertoli cells by increasing their responsiveness to FSH and simultaneously inhibits the secretion of LH by the mechanism of feed back upon the hypothalamus and the pituitary.
FSH controls the maturation of the spermatic epithelium by acting directly on the sertoli cells.
Finally the protein which binds to the androgens (ABP) is produced by the sertoli cells.
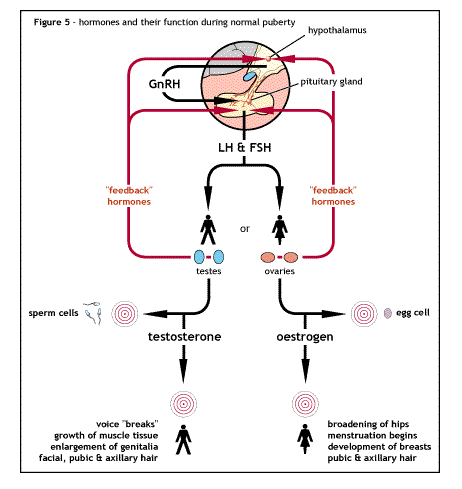 Hormonal interaction in the hypothalamus pituitary
Hormonal interaction in the hypothalamus pituitaryFSH is necessary to develop the ABP production by the sertoli cells and to develop the blood testis barrier and other functions of these cells.
Once the sertoli function is developed, testosterone alone will maintain spermatogenesis. The yield of spermatozoa is increased if FSH is present.
The FSH is known to increase the yield of spermatogonia by preventing atresia of differentiating spermatogonia.
Normally 50% of spermatogonia can also be reduced by increased sexual activity.
FSH levels in males are environmentally influenced, increased by sexual activity and decreased by inhibin.
Androgens are transported from the site of production (leydig cells) to influence the developing germ cells.
ABP produced by the sertoli cells and shed into the adluminal compartment, assists in the role as well as transporting large amount of androgens to epididymis.
First stimulates ABP synthesis under the action of androgen influence.
Testosterone induces and maintains spermatogenesis acting through the sertoli cells or through spermatogenetic cells.
The testis also secretes some other hormones that participate in the regulation of spermatogenesis, but their cells are not closely understood. These include:-
i) Estradiol formerly known as female sex hormone. These estradiol receptors are widely distributed in testicular cells, suggesting a role of oestrogens in the regulation of testicular function.
The receptors are localized in the nuclei of spermatogonia, spermatocytes and early developing spermatids of adult men.
ii) Inhibin – (Inh – b), this is produced by the sertoli cells and controls the secretion of FSH from the pituitary and consequently the spermatogenesis, via a negative feedback mechanism. Low blood concentration of inh – b of ten reflect in a disorder of spermatogenesis.
iii) Antimullerian hormone
Exclusively secreted by the sertoli cells and represents a prelocious hormonal index of their function.
Its production is influenced by transcriptional factors testosterone, FSH and spermatocytes at prophase I. It prevents formation of mullerian duct.
SUMMARY:
Mechanism of hormonal control of spermatogenesis:
The hypothalamus secretes gonadotrophin releasing hormone (GnRH) which travels in a small vein from the hypothalamus to the pituitary gland.
GnRH stimulates in turn the anterior pituitary gland to secrete two hormones know as gonadotrophins. (A gonadotrophin is a hormone that stimulates a gonad, in this case the testis). These gonadotrophins are follicle stimulating hormone (FSH) and lutenizing hormone (LH). Also secreted in female they are glycoproteins.
FSH acts by stimulating spermatogenesis by stimulating the sertoli cells to complete the development of spermatozoa from spermatids.
LH stimulates the synthesis of the hormone testosterone by the leydig cells (interstitial cells) of the testis. It is therefore known interstitial cells stimulating hormone (ICSH) in the male
Testosterone stimulates growth and development of the germinal epithelial cells (spermatogonia) to form sperms and also work with the FSH to stimulate the sertoli cells.
The negative feedback mechanism operates where by an increase in the level of testosterone results in a decrease in secretion of GnRH from the hypothalamus, this in turn results in declining levels of LH and FSH.
The testosterone also acts directly on the anterior pituitary gland to reduce LH secretion but this effect is weaker.
When the rate of spermatogenesis in high, inhibin (a glycoprotein hormone) is released, it acts on the anterior pituitary gland to reduce the secretion of FSH by negative feedback mechanism.
It also has a slight effect in the hypothalamus reducing GnRH secretion. When the rate of spermatogenesis low, inhibin is not secreted and FSH stimulates spermatogenesis.
THE ROLE OF CYCLIC AMP
Both FSH and LH acts by causing the release of cyclic AMP (Adenosine monophosphate) within the cells they stimulate.
Cyclic AMP is the second messenger system. It is released into the cytoplasm and then passes to the nucleus where it stimulates the synthesis of enzymes. In the case of LH, for example enzymes are
involved in the synthesis of testosterone from cholesterol.

STRUCTURE OF MATURE HUMAN SPERMATOZOANS Structurally, a spermatozoan is divided into three pieces:-
I) Head piece
This consists of a nucleus and small portion of cytoplasm.
At the tip of the head, there is a special structure called acrosome. Acrosome is a collection of lysosomes; it thus contains very powerful hydrolytic enzymes known as proteases and hyaluronidases.
II) Middle piece
This is largely consisting of mitochondria. These provide energy for propelling the spermatozoans towards the egg cell. The head and middle peace together constitute the principal peace.
III) The tail piece
It consists of the flagellum made of axial fillaments that continue from middle peace. The flagellum serves in:-
Propelling the spermatozoans towards the egg cell.
Orienting the spermatozoans so that it properly binds itself into the egg cell.
At the end of the flagellum is a hair like extension called the end piece.

Structure of mature human sperm ROLE OF SPERMATOZOAN.
The role is to carry the paternal gamete materials into the egg cell so that after fertilization, the genetic makeup of the zygote is the mixture of the two maternal and paternal gamete materials.
HORMONAL CONTROL OF SPERM PRODUCTION
When the level of testosterone is low, the hypothalamus secretes a releasing hormone (called gonadotrophin releasing hormone or GnRH) in the blood.
This peptide hormone flows in the blood directly to the pituitary a pre-sized organ hanging from the base of the brain where it stimulates the two peptide hormone, lutenising hormone (LH) and follicle stimulating hormone (FSH).
These hormones (called gonadotrophins, since they stimulate gonalds) then move through blood stream and activate cells in the testis. LH triggers the intestinal cells to produce and secrete testosterone.
FSH cause supporting cells (sertoli cells) to enhance formation of sperms.
Soon the sperm count rise. Mean while, testosterone circulate in the blood stream at higher levels and the interconnected loop feeds back on itself. High testosterone levels signal the hypothalamus to produce less releasing hormone.
This inturn suppress the release of LH and FSH and without them less testosterone and fewer sperms are manufactured. In addition testosterone causes supporting cells in the testes to release the peptide hormone inhibin, which helps to inhibit FSH production.
When testosterone level drops too low again the hypothalamus is once more activated and the whole cycle starts again.
ADAPTATIONS OF THE SPERMATOZOANS.
The adaptations of the spermatozoans to its function include the following:-
It has an acrosome that contains enzymes for digesting the egg cell membrane.
It has numerous mitochondria that produce energy necessary for propelling the spermatozoans towards the egg cell.
It has flagellum for propelling the spermatozoans for proper binding on the egg cells.
Ability to sense the chemical attractants secreted egg cell so that its movement is directed toward source of chemicals.
Ability to recognize and hence bind itself into the receptor sites on the surface of egg cells.
Light nuclei and head piece following their changes, this enables it to move faster towards the egg cell.
OOGENESIS
Oogenesis begins soon after fertilization as primordial germ cell travel from the york sac to the gonalds, where they to proliferate mitotically.
The gem cells multiply from only a few thousands to almost 7 million.
They become oocytes once they enter the stages of meiosis several months after birth, now called primordial germ cells surrounded by fellicle cells from the somatic line. The oocytes are then arrested in the first meiotic phase until puberty.
At puberty between 4 to 10 follicles begin to develop although only 1 – 2 are actually released.
Surrounding each oocyote is a zona pellucida membrane granule and the cell layer.
Each oocyte finishes its first meiotic division creating a secondary oocyte and polar body which serves no further functions.
It begins the next meiotic cycle and is arrested in its second metaphase, at which point it is released from the ovary in ovulation.
It will not finish the meiosis cycle until it encounters the stimuli of a sperm.
SUMMARY:
Oogenesis
At birth.
Diploid cells in the ovary divide by mitosis from oogonia.
Oogonia undergo meiosis I division to form primary oocyte steps at prophase I.
These remain in the follicles.
At puberty.
Primary oocyte completes the 1st complete meiotic division to form polar bodies and secondary oocyte.
Secondary oocytes undergo 2nd meiotic division and steps at metaphase II.
Secondary oocyte is shed from ovary, if fertilized it complete its 2nd meiotic division to form ovum and polar bodies.
Formation of the ovum involves substantial increase in cell volume as well as the acquisition of organelles that adapt the egg for reception of the sperm nucleus and support of the early embryo.
In the fetal ovary of mammals the oogonia undergoes meiotic divisions until the birth of the foetus, but the process involves the destruction of the majority of the developing ova by the seventh month of gestation reduces the number of oocytes from millions to a few hundred around the time of birth, the mitotic divisions ceases altogether and the fast female contains its full complement of potential ova.
|
Weak of gestation
|
Stages
|
No of germ cells
|
|
¾
|
Primordial cells in the endoderm of the yolk sac.
|
|
|
5 – 6
|
Premeiotic cells oogonia.
|
10,000
|
|
8
|
Propagation by mitosis.
|
500,000
|
|
8 -20
|
Mitosis, meiosis atresia maximum at week 20.
|
6 – 700,000
|
|
20 -40
|
Reduction of oocyte, 80% of germ cells are lost.
|
1-2,000,000
|
|
Birth to puberty
|
Further oocytes are lost by atresia.
|
|
Unlike the formation of sperm in which the two divisions of meiosis produce four equivalent daughter cells, the cytoplasm of the oocyte is divided unequally so that three polar bodies with reduced cytoplasm and one oocyte are final products.
Egg cytoplasm contains large stores of ribonucleic acid RNA in the form of ribosomal messenger and transfer RNA. These RNA’s direct the synthesis of proteins in the early embryo and have decisive influence on the course of development.
DEVELOPMENT OF GERM CELLS IN THE OVARY
Following the immigration of the primordial germ cells into the gonadal ridge, they proliferate are enveloped by coelomic epithelial cells and form germinal cords that though keep their connection with the
coelom epithelium.
In the genital primodium, the following processes take place;
A wave of proliferation begins that lasts from 15th weak to the 7th month.
Primary germ cells arise in the cortical zone via mitosis of oogonia dones, bound together in cellular bridges that happen in rapid succession.
The cell bridges are necessary for a synchronous onset of the subsequent meiosis.
With the onset of meiosis earliest in the prophase in the 12th week the designation of the germ cells change. They are now called primary oocyte.
The primary oocyte become arrested in the diplotene stage of prophase I the prophase of the 1st meiotic division.
Shortly before birth, all the total oocytes in female ovary have attained this stage.
The meiotic resting phase that then begins is called the dictyotene and it lasts till puberty during which each month and each month thereafter until menopause a pair of primary oocyte complete the first meiosis.
Only few oocytes, secondary oocyte plus one polar body though reach the 2nd meiosis and the subsequent ovulation. The remaining oocytes that mature each month become atretic.
The primary oocytes that remain in the ovaries stay in the dictyotene stages up to menopause. In extreme cases without ever maturing during the menstrual cycle.
From birth, there are thus two different structures to be distinguished that at least conceptually do not develop further synchronously.
Female germ called primary oocyte and which can develop further only during and after puberty hormonal cycle is necessary.
Follicular epithelium, that can develop further from the primordial follicle via several follicle stages while oocytes remain in their primary states.
The developmental sequence of the female germ cell is as follows:-
 The continous of the development/maturation of the oocyte begins again only a few days before ovulation.
The continous of the development/maturation of the oocyte begins again only a few days before ovulation.The developmental sequence of a follicle goes through various follicle stages.
Since the follicle can die at any moment in this development (atresia) not all reach the tertiary follicle stage.
PRIMARY OOCYTE
In the first week of the cycle the maturation of the oocyte in its associated follicle depends on the progress of maturation of the surrounding follicle cell.
The fittest follicle with its oocyte becomes the dominant follicle in the second cycle week and later the graafian follicle.
Up to just days before ovulation, the maturation of the oocyte consists in its ingestion of substances growth of the yolk, they are supplied by the granulose cells. This exchange of substances is mediated through cytoplast processes of the granulose cells that are anchored through zona pellucida at the ocoyte substance.
The oocyte nucleus is also matured in the last days before the LH peak.
Up to this point it was arrested in the extremely elongated prophase (a dictyotene) of the first meiotic division, the corrested condition that has existed since the foetal period.
Through the maturation the nucleus stages in the darkness of the prophase and prepares itself for the completion of the first meiosis which is triggered by the LH peak.
With the LH peak, the following maturation steps are now triggered in and around the oocyte up to ovulation.
In the oocyte:
Termination of the first meiosis with ejection of the first polar body.
Begin of the 2nd meiosis with arrest in the metaphase.
Maturation of the oocyte cytoplasm by preparing molecules and structures that will be needed at the time of fertilization.
In the follicle:
The granulosa cells that sit just outside on the zona pellucida withdraw their processes from oocyte surface back into the pellucida zona. These processes were in charge of transferring substances to the oocyte.
The periritelline space forms between the oocyte and the pellucida zona. This space is necessary for allowing division of the oocyte and for harbouring the first polar body formed in the division.
Loosening of the granulosa cells in the vicinity of the cumulus oophorus and proliferation of the granulosa cells.
Increasing the progesterone concentration in the follicle fluid via increased production in the granulosa cells.
Termination of the first meiosis
The spindle apparatus for dividing the chromosomes has formed and oriented itself radically in the cellular surface.
The first polar body will arise at the spot where the spindle apparatus is anchored on the cellular surface.
The process of the granulosa cells have refracted from the oocyte surface into the pellucida zona and this leads to the formation of the pervitelline space, in this space the ejection of the 1st polar body takes place as a sign that the 1st meiosis has ended.
With the end of the first meiosis the name of the changes from primary oocyte to secondary oocyte.
The secondary oocyte
Through the effect of LH on the granulosa cells, these have begun to loose their cellular bends and to multiply.
They produce progesterone that is released into the fellicle fluid.
Though the separation of the homologous chromosomes in the first meiosis a haploid (reduplicate) set of chromosomes is now to be found in the secondary oocyte.
The role of progesterone in the follicle fluid
Progesterone has the following two main tasks in the follicle fluid:-
It stimulates the further maturation of the oocyte.
During ovulation, it enters the fallopian tubes and guides the formation of a concentration gradient for attracting the sperm cells.
The follicle that is about to rapture:-
Besides the hormones the granulosa cells also secrete an extra – cellular matrix, mainly hyaluronic acid, into the follicle fluid.
The cumulus cell bonds loosen further in this way together with the enclosed oocyte they free themselves from where they were attached to the follicular wall and in the follicle.
The wreath of granulosa cells that enclose the oocyte is called the corona radiate.
The oocyte has now ended all the steps of maturation that were set into motion by the LH peak.
The molecular and structural preparations for the time following the penetration by the sperm cell have now been made in the cytoplasm.
A spindle apparatus (2nd meiosis) has again been able to form with the chromosome in the equational level (metaphase plate)
The spindle is once more anchored radially to the cell membrane near the polar body.
The same processes of the spindle formation also take place in the polar body.
NB: The second meiosis is arrested in this position.
Final steps of the maturation namely the freezing for the second meiosis are first completed by the second oocyte when the spermatozoa have penetrated the oocyte.
The follicle and the oocyte are now ready for ovulation that takes place roughly 38 hrs after the LH peak.
ADAPTATION OF THE EGG CELL
Has microvilli for nutrient absorption from follicular cells.
Follicular cells are the cells that usually surround the ovum when more layers are formed they tend to push away the follicular cells.
NB: The follicular cells are not part of the egg cell.
It has stored food for zygote and embryo utilization (i.e. the yolk sac).

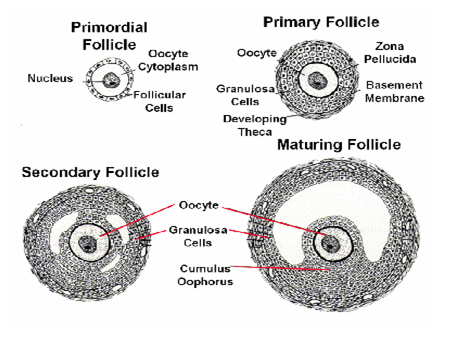
Fig: The egg cellFig: The stage of oogenesis
Has cortical granules (Act as vesicles) that act to prevent polyspermy during fertilization.
This is done through two ways:-
The process occurs very quickly and uses the same mechanism as in a chemical transmission of impulses. It occurs soon after the entry of a sperm cortical, are found in the cytoplasm.
They fuse with the membrane to release chemicals which haidens the membrane forming the fertilization membrane that prevents the entry of sperms.
Destruction of the sperm receptor sites. The sperms have sensors and ova have receptors-sperms move towards the ovum (like magnetic substance) chemotactically. The sperms receptor sites found on the ovum are destroyed immediately after the entry of a sperm. This is done by the cortical granules.
2.Have receptor sites for spermatozoa to bind during fertilization.
3.Produce chemicals that attract sperms.
DIFFERENCES
|
SPERMATOZOA
|
EGG CELL
|
|
Small in size.
|
Larger than a sperm.
|
|
Has a large nucleus.
|
Has a smaller nucleus.
|
|
Has a very small amount of cytoplasm.
|
Has a very large amount of a cytoplasm.
|
|
No food reserves (does not store food).
|
Stores large amount of food.
|
|
Has acrosome.
|
Has no acrosome.
|
|
No cortical granules.
|
Has cortical granules.
|
|
Has head, middle piece principal and end piece.
|
No such division.
|
|
No microvilli.
|
Has microvilli.
|
|
Single layered.
|
Multi layered.
|
|
It has flagellated and motile.
|
It lacks flagellum and non motile.
|
|
It has numerous mitochondria.
|
Has few mitochondria.
|
DIFFERENCE BETWEEN OOGENESIS AND SPERMATOGENESIS
|
SPERMATOGENESIS
|
OOGENESIS
|
|
Differentiation follow offer its meiotic division they are farmed only until the end of meiosis.
|
– Egg grows primarily in extend period of prophase i.e. prophase that is secondary oocyte is already matured.
|
|
It occurs in male gonads i.e. testis.
|
– It occurs in female gonad i.e. ovaries.
|
|
Four sperms are produced from one spermatogonium.
|
– Only one ovum is produced from one oogonium.
|
|
The spermatocyte sperm mother cell divides by meiotic division into four equal sized cells and all the four cells are transformed into spermatozoa to act as reproductive unit.
|
-The oocyte divides unequally and produces a large sized ovum and three small sized bodies or polocytes which are sexually inert only ovum acts as reproductive unit.
|
|
Spermatozoa are produced in large number.
|
-Ova are produced in large number.
|
|
Spermatozoa are minute yolkless and motile.
|
– Ova are much large, often with yolk and non-motile.
|
|
Continous production process although from puberty to old age sperm cells are being endangered, the production is subject to extreme fluctuations regalding both quality and quantity.
|
– Using up the oocyte generated before birth, continual decrease of the oocyte, beginning with the foetal period-exhaustion of supply at menopause.
|
|
During foetal period no meiotic division, no germ cell production.
|
– During focal period, entering meiosis (arrested in dictyotene stage), there is a production of entire supply of germ cells.
|
FERTILIZATION
Is the process whereby the nucleus of the male gamete fuses with that of the female gamete to form diploid zygote nucleus.
This process occurs high up in the fallopian tube. Before fertilization the spermatozoa has to undergo capacitation.
CAPACITATION
Is the mechanism by which the spermatozoa undergo activation before fertilizing the ovum. It takes about 7 hours and involves the following processes.
Removal of a layer of glycoproteins and plasma proteins from the outer layer/surface of spermatozoa; glycoprotein are added by the epididymis while the plasma proteins are from the semen. These are removed by the enzymes in the uterus.
Removal of cholesterol which toughens the sperm membrane and prevents premature release of acrosomal enzymes from the sperm head membrane by the enzyme in the uterus.
The advantage of capacitation is that it prevents the wastage of sperms. The membrane becomes more permeable to Ca2+ ions.
The calcium ions (ca2+) have two factions:-
Increase/enable the beating of the sperm flagellum.
Promote acrosomal reaction.
MECHANISM OF FERTILIZATION
Fertilization comprises of two types of chemical reactions:-
Acrosomal reactions.
Cortical reactions.
STEPS:
The sperm migrates through the coat of follicle cells and binds to a receptor molecule in the zona pellucida of the egg.
The binding includes the acrosomal reactions in which the sperm releases digestive enzymes into the zona pellucida. These enzymes are proteases (acrasome) and hyalurionidase. The latter digests the hyaluronic acid which binds granulosa cells together.
With the help of this hydrolytic enzyme, the sperm reaches the egg and the membrane protein of the sperm binds to the receptor on the egg membrane.
This induces the influx of Ca+ ions that depolarize the egg membrane. This is the first block of polyspermy.
The plasma membrane fuses making it possible for sperm nucleus to enter the egg.
The sperm egg cell fusion causes Ca2+ influx.
This inturn triggers a cortical reaction in which secretions beneath zona pellucida. These secretions swell up with water, push any remaining sperms away from the egg and creating impermeable fertilization membrane. Then the enzymes harden the zona pellucida. This functions as the flow block to polyspermy.
The nucleus of the secondary oocyte is triggered to undergo meiosis II which produces an octid is transformed into an ovum. The nucleus of the ovum and that of spermatozoa bulge becoming pronida which later fuse in the actual act of fertilization forming zygote.
NB:
If not fertilized, the secondary oocyte dies off ovulation and never finishes meiosis.
POST – FERTILIZATION CHANGES IN THE EGG
After fertilization, the following changes occur in the egg.
The zygote becomes ready for the cleavage and for the formation of the embryo.
The oxygen consumption of the zygote increases enormously.
The metabolic rate of the zygote increases greatly for instance the amount of amino acids and the permeability of the plasmalema of the egg increases the volume of the egg decrease the exchange of phosphate and sodium ions between the zygote and the surrounding media, diffusion of the calcium ions from the egg started and the hydrolysing activities of the
Protein synthesis in started.
SIGNIFICANCE OF FERTILIZATION
The fertilization ensures the usual specific diploid of the organisms by the fusion of the male and female pronuclei.
The fertilization establishes definite polarity in the eggs. This fertilization provides new genetic constitution to the zygote.
The fertilization activates the egg for the cleavage.
Fertilization provides a new genetic constitution to the zygote.
Fertilization combines characters or two parents thus introducing variations and making the resulting individual better equipped for the struggle for existence. This happens only in cross fertilization.
The fertilization also increases the metabolic activities and the rate of protein synthesis of the cell.
THE CONCEPT OF STERILITY
STERILITY:
Is the failure of the matured mammal to fertilize or to be fertilized.
CAUSES OF INFERTILITY
FEMALE INFERTILITY MALE STERILITY/INFERTILITY
|
i) Failure to ovulate due to hormonal causes.
|
– Absence of sperm due to blockage of tubes between the testes and seminal vesicles.
|
|
ii)Uterus damage, pregnancy cannot be maintained (occurrence of miscarriage).
|
– Low sperm count.
|
|
iii)Damage to the oviduct due to tubal diseases hence oviduct is blocked.
|
– Production of abnormal sperm.
|
|
iv) Cervix damage due to abortion or difficult birth hence inability to produce cervical mucus for sperm to reach the egg.
|
– Autoimmunity: Antibodies are made by the male body which attract the sperm reducing sperm count.
|
|
v)Antibodies to sperm.
|
– Impotence.
|
IMPOTENCE:
Is the failure of penis to erect, this can be temporary i.e. reversible impotence caused by such factors as;-
Depression due to social, economic and ethnic reasons.
Fear due to inferiority complexity, disease contraction, hesitating to commit sin.
Also impotence can be permanent irreversible due to genetic disorder, diseases, hormonal problems etc.
Copulation – (i.e. seduction, romance and the subsequent intercourse) ensures the transfer of sperms from the male reproductive organs to female reproductive organs for fertilization.
DEVELOPMENT OF THE ZYGOTE AND EMBRYO
This includes 5 stages:
1)Cleavage
2)Blastulation
3)Gastrulating
4)Neurilation
5)Organogenesis
CLEAVAGE
Two hours after fertilization the zygote divides mitotically to form two cells.
The process by which the zygote divides is called cleavage and the resulting cells are called blastomeres.
After 6 hours the zygote cleaves for the second time forming four blastomeres.
Initially the process is regular but with time it becomes irregular in where it produces a ball of cells called MORULA.
Cleavage does not lead to increase in size of the morula because cells still in the zona pellucida.
The process takes place 72 hours.
Cleavage increases surface area to volume ratio of each cell which enhances:-
Rapid nutrient uptake i.e. food and oxygen.
Waste removal.
Cleavage also forms many cells which will form different types of embryonic tissues.
BLASTULATION
Is the process whereby morula is transformed into a blastula or blastula or blastocyst.
While cleavage is taking place the zygote is in the oviduct moving slowly by the beating action of the cilia in the oviduct (tubules). When it reaches in the uterus the hard zona pellucida gets peeled off by the enzymes in the uterus and leaves an outer layer of cells called trophoblast.
The cells in the centre of the morula migrate and accumulate at one end where they form an inner cell mass; the result of this cellular migration is the formation of the fluid filled cavity called bastocoel.
IMPLANTATION
Is the process whereby the blastocyst embades into the uterine wall.
As soon as the trophoblast is in contact with the uterine wall it starts secreting enzymes that eat through the endometrium wall thereby pouring a way for blastocyst to embed.
Trophoblast develop finger like processes called trophoblast villi (chorion villi) which are for the absorption of nutrients from the uterine wall. The trophoblast is also endocrine in function as it secretes a human chorionic gonadotrophs hormone (HCG).
The function of the HCG is like that of LH.
To maintain the corpus luteum secretion of oestrogen and progesterone also done.
Inhibit menstruation to pregnant woman.
Forms the basis of the pregnancy test (urine – pregnancy test UPT) dip the litmus paper into the urine if you see two red marks; the person is pregnant; if only one colour is seen then the person is not pregnant.
The process of penetrating in the uterine wall continues until finally the blastocyst becomes completely embeded in the glands and blood vessels of the uterus. This is the actual act of implantation.
GASTRULATION
This is transformation of blastula into germ layer called gastrula.
It s a stage at which the embryo develops germ layer.
During gastrulation, the cells on one side of the embryo inviginates forming a small pore called blastopore.
The process ends when germ layers are ready formed.
NB: All the five stages are summarized in the diagram below:-
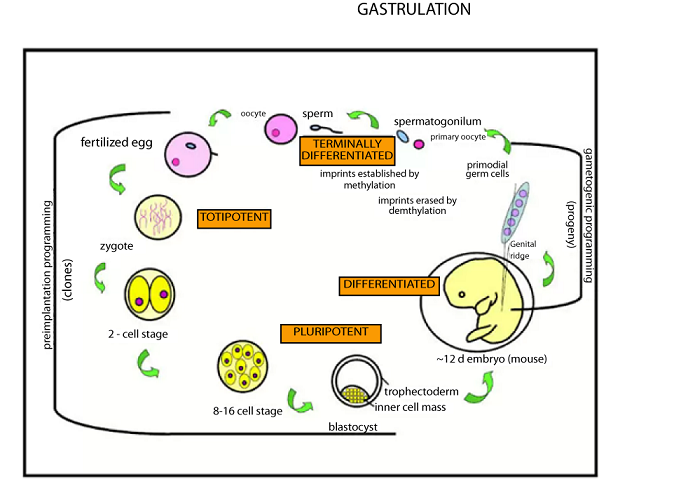
Through this pore, blastopore about half of the cell from outside move to the inside and at this point, the embryo is said to turn on itself.
The result of this cellular migration is the development of two germ layers, the outer (ectoderm) and the inner (endoderm).
The blastocoel becomes an archenteron the future digestive tract. The blastopore is the future anus.
Finally the third layers the mesoderm form between the ectoderm and endoderm forming a three layered embryo.
Gastrulation is important in placenta development because cell’s location in a particular layer determines its fate e.g.
Ectoderm – Develops nervous system, sense organs, epidermis the skin, hair nails and skin glands, neural egest.
Mesoderm – Develops into bones blood, muscles, dermis of the skin and reproduction system.
Endoderm – Develops digestive and respiratory system and many glands.
EXTRA EMBRYONIC MEMBRANES AND THEIR ROLES
After implantation the embryo develops four membranes. They are called extra embryonic membranes because they are found outside the embryo and these include:-
i) Chorion
ii) Amnion.
iii) Allantos.
iv) York sac.
I. CHORION
This is the outer most membrane which is derived from the trophoblastic cells.
It has villi that forms the part of the placenta, therefore the roles of chorion are;-
To form parts of the placenta.
To absorb nutrients from the mother to the foetus means of villi.
Since it is an outer member, it then protects the foetus.
II. AMNION
This is the innermost membrane which lines the cavity surrounding the embryo.
This cavity (amniotic cavity) is filled with the amniotic fluid secreted by the amniotic cells.
The amniotic fluid acts as shock absorber cushioning the embryo against mechanical and physical shock.
III. ALLANTOIS
This is a sack like outgrowth which develops from embryonic gut; it fuses with the chorion at the point called allantois-chorion where the placenta develops.
As the embryo continues to grow the allantois develops into umbilical cord the tube which carries blood vessels (embryonic) to end from the chorionic villi.
IV. YOLK SAC
This has got no obvious function in humans and other mammal it becomes buried in placenta.
In reptiles and birds the yolk sac is important as it absorbs food from the yolk and transfers it to the midgut of the developing embryo.
PLACENTA
A placenta begins when extension of chorionic villi penetrates more and more deeply into the endometrium like the roots of a tree in the ‘soil’ uterus.
As they digest their way through the uterine blood vessels the villi become surrounded by pools of free blood the latter forms placental sinuses.
A placenta is a linking structure between the foetus and the mother. It is the structure that partly develops from the mother and parity develops from the embryo. It thus has the foetal and maternal side.
At the placenta, the materials are exchanged between the foetus and the mother. However, their vascular systems are not in physical contact. The exchange of materials is therefore by simple diffusion.
Why are the material and foetal blood not allowed to mix?
Maternal blood is under relatively higher pressure compared to foetal blood; this could damage the delicate tissues of the developing foetus
If the two bloods were to mix, the foetal blood could be recognised as a foreign by maternal blood. The maternal blood immune system could respond by killing the foetus.
This is because half of the genetic materials come from the father and so the foetal cells are not identical to those of the mother.
i) Progesterone.
ii) Oestrogen.
iii) Human chorionic gonadotrophic hormone.
ROLES OF PLACENTA
It allows the exchange of materials between the foetus and the mother without mixing up the two blood.
It is a means of passage of oxygen, water, acids, glucose (i.e. nutrients) to the foetus (acts as intestine).
Means of passage of carbondioxide, urea and other wastes from the foetus to the mother so as to allow the excretion by the mother and prevent harmful substances to accumulate in the foetus i.e. acts as lungs and kidneys.
Allows certain antibodies to pass into the foetus providing it with some immunity against diseases. This is called Natural passive immunity.
It protects the foetus by preventing certain pathogens and their toxins from crossing the placenta. Though, some manage to cross. Eg. Treponema pallidum (for syphilis) and HIV.
It prevents hormones and some chemical substances like alcohol to pass through the foetus.
Qn:
Placenta serves as a link between foetus and mother. At the same time it acts as a barrier between them. By reference to the functions of placenta explain what those statements mean.
|
Placenta as a link
|
Placenta as a barrier
|
|
O2, H2O, food, salt from the mother to the foetus.
|
Prevent blood mixing.
CO2 and other nitrogenous wastes from foetus to mother.
Prevent high maternal blood affecting foetus directly.
|
|
Partly antibodies from the mother to the foetus.
Filter out some hormones
Partly filter out some pathogens.
|
|
TWINS PUZZLE AND MULTIPLE BIRTH
MULTIPLE BIRTH AND THEIR CAUSES:
Multiple birth are cases in which more than one baby are born from the same mother and they result from the same pregnancy.
In mammals like cats, rabbits, dogs and pigs multiple birth are common cases as the ovulation several oocytes are released each of them is fertilized by separate spermatozoan.
Humans are commonly giving birth to only one young individual.
Multiple birth occurs due to;
More than one secondary oocyte being released at ovulation and then fertilized by spermatozoa.
One ovum being fertilized by spermatozoan and zygote cleave into 2,3 …etc blastomeres each of which develops into an embryo after separation.
TWINS
Defn: Are two or more babies born from the same mother as a result of the same pregnancy.
TYPES OF TWINS:
Identical twins
Result from the same zygote/one zygote hence called monozygotic twins. For the development of identical twins to occur the zygote cleaves into two or more blastomeres.
These separate from one another and upon implantation each one of them develops into an embryo.
In rare cases, separation of the blastomere fails at some points. Thus leads to twins remaining linked, such twins are referred to as SIAMESE TWINS.
The identical twins share the same placenta and they are in the same chorion and amnion.
Since they develop from the same zygote such twins are of the same genetic constitution and of the same sex.
2. Flatenal/Non identical twins
They develop from two different zygotes hence they are called dyzyotic twins.
In this case two different ova from different ovaries are fertilized by two different spermatozoa forming two different zygotes, which implant in the uterus.
Each of these twins develops in its own placenta and its own embryonic membranes since they develop from different zygotes. Then the zygotes are genetically different and not necessary of the same sex.
DIFFERENCES
|
IDENTICAL TWINS
|
NON- IDENTICAL TWINS
|
|
They result from one zygote.
|
Results from two different zygotes.
|
|
Share the same placenta.
|
Each has its own placenta.
|
|
Enclosed in the same membrane.
|
Each has its own membrane.
|
|
They are of the same genetic makeup.
|
Genetically different.
|
|
They are of the same sex.
|
They may be of different sexes and can be a girl/boy.
|
BIRTH(PARTUIRITION)
Birth is a process whereby the fully developed features expelled out of the mother’s womb after the GESTATION period is complete.
THE PROCESS OF BIRTH/LABOUR
The labour occurs in three stages shown by distinct events. These events are longer in primigravide than in multigravide.
THE FIRST STAGE
This is the stage of labour pains. During this stage the fully developed foetus has its own hypothalamus stimulated to release ACTRF which in turn stimulates the release of ACTH from the foetal pituitary gland.
The ACTH stimulates foetal gland (adrenal) to release cortical steroids. The released cortico steroids pass across the placenta and enter the maternal circulatory system where they perform the following:-
I) They cause increase in prostaglandins (secreted by uterus).
II) They cause decrease in progesterone following the decrease in progesterone.
The pituitary gland is allowed to release oxytocin.
The inhibitory effect on myometrium contraction is removed and prostaglandins power the contraction.
Oxytocin therefore causes the contractions of the myometrium where as the prostaglandins secreted by uterus increase the power of contraction. These contractions of myometrium sum up to labour pains.
As the uterine walls continue to contract, the cervix dilates under the influence of hormone called relaxin. The amnion and chorion rupture releasing the amniotic fluid through the cervix breaking of water. Contractions continue from top to bottom forcing the baby out of the womb.
The body gets engaged into the pelvis and following further contraction, the foetal head gets into the cervix where it causes irritation and increases powers of contraction.
The first stage of labour is terminated .When the diameter of the head is equal to the diameter of the cervix.
THE SECOND STAGE
Is the stage where by the baby is completely delivered out of the mother’s womb.
As soon as the baby is out, the umbilical cord is ligatured at two points and a cut is made between the two ligatures so as to make the baby totally separated from the mother’s physiological reliance.
THE THIRD STAGE
This is the stage during which the placenta and the extra embryonic membranes after birth is delivered.
The birth of after birth is due to dramatic contraction of the uterus which causes the placenta to detach from the uterine wall.
It is important that after birth is not allowed to remain inside for a long time as its decomposition leads to blood passing.
PARENTAL CARE
This comprises of all activities the parents do for the better growth and bringing of their offspring or the offspring of the near relative.
ASPECTS OF PARENTAL CARE
Nutrition -3 month
The body has to be fed on the nutritious food substances, for proper growth and development. After birth, the baby is fed on breast with from its mother.
Breast feeding is highly recommended because mother’s milk contains all important food substances needed by the baby at every stage of its development.
The first milk that a baby is sucking from the mother’s breasts is a special one as it is called colustrum.
This is a yellowish fluid that contains antibodies to provide immunity to the baby. After the first three months, the child continues to be supplied with extra proteinous food substances such as cow’s milk,
eggs, fish, beef etc. for proper growth and development.
Protection
Most of the mammalian parents protect their young against
a) Disease: by providing health services ensuring hygienic handing of food etc.
b) Climate changes: by providing warmth to the babies.
c) Predators/enemies: mothers become agressive to ensure that their young ones are not reached.
2. Social interaction/Education
The young ones must learn how to interact with others and fit into the social structure around them. The youngs need early experience with their parents in order to depend on themselves and to learn
to live actively in the social unit.
Parental care involves the:-
Learning of language.
Teaching the language.
Formal education (For human beings, primary, secondary up to tertiary education) thereafter a person becomes independent.
REPRODUCTIVE CYCLES
In order to syncronise the favourable conditions to sexual reproduction, sexual reproduction is naturally in cycle.
For example; plant flowering is at the same date year after year; bleeding (menstruation) of a mature primate is on the same date month after month etc.
In mature female mammals, there is a sexual reproduction cycle, this is known as oestrus cycle (ovarian cycle).
At the onset of puberty there are approximately 400,000 primordial follicles and single follicles in all stages of maturity in the ovary. Ooctyes contained in the primordial follicles migrate out of the extragenital
structures of the coelomate epithelium into the stroma of the primary bipotent gonalds as oogonia during embryonic development.
These then divide mitotically of the roughly 400,000 follicles that are present in the two ovaries at the beginning of sexual maturity, only around 480 reach the graafian follicle stage and are thus able to
release oocytes (ovulation).
This number is simply derived by multiplying the number of oocytes of cycles per year (12) and the number of years in which a woman is fertile (40).
Cyclic ovarian functions entailing follicle formation, ovulation, corpus luteum development and luteolysis is regulated by the hypothalamus pituitary system as well as by intraovarian mechanisms
hypothalamus, pituitary and ovary are there by in dynamic interaction.
OESTRUS CYCLE
Defn:
Oestrus cycle is the total time taken for the development and degeneration of an ovarian follicle.
In some mammals, this period occurs once in a year, they are said to be monoestrus eg. fox.
Note that; menstrual cycle in human, oestrus cycle in other mammals.
The discharge of the blood marks the end of oestrus cycle in higher mammals of order primate.
In most mammals, this period is occurring many times in a year, so they are polyestrus.
PHASES OF OESTRUS CYCLE(VERY MINOR)
Anoestrus – Period during which no visible sexual activity in females.
Proestrus – Period during which graafian follicle develops into ovary and secrete oestrogens. Also called follicular phase.
Oestrus (heat) – Ovulation normally occurs, the female is ready to mate and becomes sexually attractive to male.
Met oestrus (luteal phase) – corpus luteum develops from raptured follicle.
Dioestrus – Progesterone secreted by corpus luteum prepares uterus for implantation.
SIGNIFICANCE OF OESTRUS CYCLE
Since it is characterised by ovulation and hence increase sexual urge of female, then it is important in that copulation is syncronised with fertilisation.
MENSTRUAL CYCLE
This approximately monthly cycle of events associated with ovulation that replaces the oestrus cycle in most primates i.e. human chimpanzee, gorilla, baboon etc.
The lining of the uterus becomes progressively thicken with more blood vessels in preparation for implantation of a fertilized egg (blastocyst).
Ovulation occurs during the middle of the cycle (fertile period). If fertilization does not occur the uterine lining breaks down and discharged is known as period.
In women the fertile period is 11 – 15 days after the end of the last menstruation.
EVENTS OF MENSTRUAL CYCLE/OESTRUS CYCLE
Day 1 and 2 anterior pituitary gland releases. FSH and LH.
These hormones travel by blood and reach the ovary.
They stimulate the granulosa cells to secrete oestrogen.
Oestrogen thickens endometrium and inhibits FSH and LH.
Day 12 LH level rises.
It stimulates granulosa cells to stop producing oestrogen and start release progesterone.
Causes ovulation.
At day 14, secondary oocyte bursts out of the ovary.
Granulosa cells filled with yellow substance to form corpus luteum.
Corpus luteum secretes progesterone.
Endometrium thickens.
Inhibits FSH and LH.
Drop in FSH and LH stops progesterone and oestrogen level going up because granulosa cells are no longer stimulated.
Endometrium no longer thickens and lack of progesterone and oestrogen stimulate the anterior pituitary gland to release FSH and LH.
Cycle begins again.
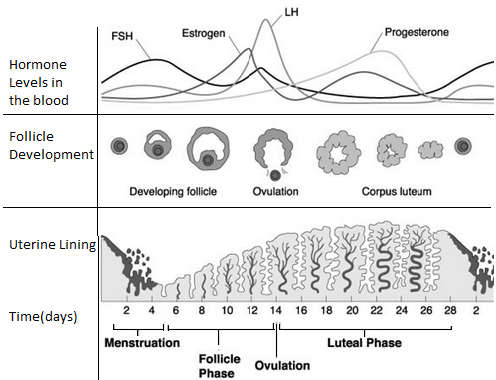
The figure above shows morphological and endocrinological changes during various phases of the cycle.
PHASES OF MENSTRUAL CYLE
1.FOLLICULAR PHASE
This is characterised by:-
Increased TSH from pituitary gland.
Production of LH from pituitary gland.
Development of follicle.
2.OVULATION
This involves the release of secondary oocyte after maturation of graafian follicle. This process in controlled by LH. One follicle rapidly out places the others and attains a diameter of up to 2.5 cm. This follicle is called mature vesicular (graafian follicle).
It is produced from the surface of the ovary like a bluster. As it develops the primary oocyte completes meiosis I producing a secondary oocyte. This begins meiosis II.
3.LUTEAL PHASE
Development of corpus luteum following ovulation; the rapture of graafian follicles develops into a yellow body mass called corpus luteum. The latter is endocrine in function and thus it secretes two hormones. Progesterone (large amount) and oestrogen (small amount).
4.MENSTRUATION
This is characterised by withdrawal of progesterone following the regression of corpus luteum also discharge of blood from vagina.
EVENTS OF MENSTRUAL CYLCLE
The events of menstrual cycle involves:-
Ovarian cycle – ovaries.
Uterine cycle – uterus.
In women, the rythmic hormonal influence leads to the following cyclic events.
The ovarian cycle (follicle maturation) that peaks in the ovulation and the subsequent luteinization of the granulosa cells.
Cyclic alternation of the endemetrium that prepare the uterine mucosa to fertilized oocyte (as ‘nest’ thene).
THE OVARIAN CYCLE
A rule, the ovarian cycle lasts 28 days (in majority).
It is divided into two phases:
i. Follicle phase: Requirement of a so called follicle and within this selection of the mature follicle. This phase ends with ovulation. Oestrogen (estradiol) is the steering hormone normally it last 14 days but this varies considerably.
ii. Luteal phase: Progesterone production by corpus luteum.
The control circuit of the hormonal cycle has two essential elements:-
i. The pulsative liberation of GnRH as well as FSH and LH.
ii. The long – loop feedback effect of oestrogen and progesterone on the hypothalamic hypophysical system.
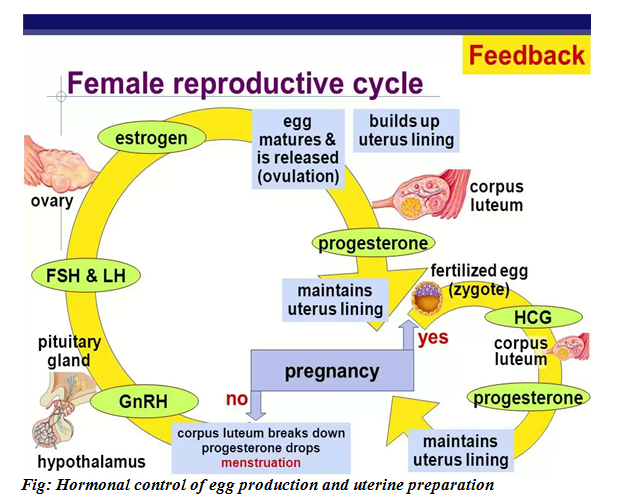
HORMONAL CONTROL OF EGG PRODUCTION AND UTERINE PREPARATION
When levels of progesterone and oestrogen in the blood are low, the hypothalamus is triggered to secrete releasing hormones (GnRH).
The releasing hormone stimulates the pituitary gland to produce FSH and LH which travel in the blood stream to the ovary.
The FSH stimulates follicles to grow but usually only one follicle with its oocyte mature each month.
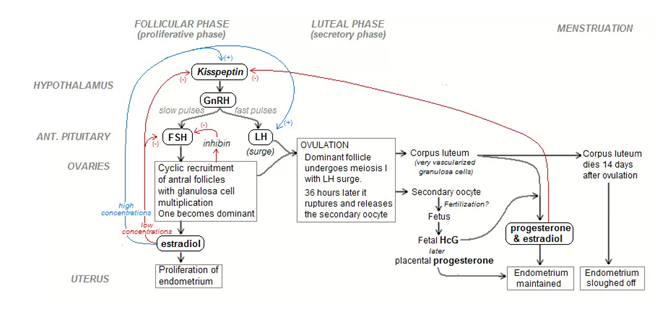
Fg: Interaction between hypothalamus pituitary gland and ovary representation of -ve and +ve feedback mechanisms.
The follicle grows rapidly and secretes increasing amount of oestrogen.
This hormone oestrogen causes the uterine lining to become thicker and more heavily supplied with blood.
On about the 14th day of a 28 days cycle the pituitary gland secretes a large pulse of LH and additional FSH and these trigger the oocyte to complete the first meiotic division it began before birth.
The developing follicle then rapture and releases the egg.
Once the ovum has left the ovary and begins its trek down the oviduct, the follicular cells left behind in the ovary enlarge and form a new gland the corpus luteum (literally yellow body).
The corpus luteum cells continue to secrete oestrogen but they begin now for the production of large quantities of progesterone as well.
Together oestrogen and progesterone promote the continual build up of the uterine lining.
The hormones as well inhibit the hypothalamus from making releasing factors and the pituitary from releasing FSH and LH.
If the ovum does not encounter sperm on its down ward journey and it is therefore not fertilized diminishing levels of LH and FSH allows the corpus luteum to degenerate on day 24 of a cycle.
Corpus luteum thus releases less and less oestrogen and progesterone.
As these hormones diminish; the endometrium begins to slough off and an approximately five days long period of menstrual flow starts making the beginning of the next cycle.
Fig: Hormonal control of egg production and uterine preparation
The control of the circuit of the hormonal cycle:
The control begins in the hypothalamus which produces gonadotrophin releasing hormone.
The GnRH is received by receptors in the anterior pituitary gland which responds by releasing follicle stimulating hormone (FSH) and lutenizing hormone (LH) in a pulsatile manner.
At the beginning of the development, the granulosa cells express FSH receptors which stimulate growth of the follicle. Theca cells express receptor for LH which stimulates the growth of corpus luteum.
Theca cells also produce andogens which the granulosa cells convert into oestrogen.
Oestrogen acts back on the anterior pituitary gland to further FSH and LH surges, and also supports the growth of the endometrium.
At some point the dominate follicles begins to secrete inhibin, which acts back on anterior pituitary gland to stop producing FSH. Only the dominant follicle which is now FSH independent will continue to grow.
During further growth/development; the granulosa cells increase their FSH receptors and express LH while the theca cells increase LH receptors.
This surge in hormone receptor results in ovulation.
After ovulation, if fertilization occurs; the corpus secretes progesterone that supports the further growth of endometrium. If, however fertilization does not occur, then the hormone level drops the corpus luteum breaks down, no longer secrete progesterone, so that the endometrium sloughs off producing menstruation.
It is estimated that lens than 1% of all follicles reach the stage of the graafian follicle with 99% of follicles degenerating by apoptosis programmed cells death is an energy dependent process accompanied by DNA degeneration.
The corpus luteum develops out of the raptured follicle immediately following ovulation corpus luteum is a vascularized version of the previous a vascular follicular epithelium with its intergration into the circulatory system and the expression of the low density lipoprotein receptors, the follicular epithelial cells are able to take up cholesterol from the periphery and use it for progesterone biosynthesis.
NB:
The origin of germ cells (gametes) is of special interest because the differentiation of these cells is responsible for continuing life cycle. The initial determination of cells as premordial germ cells occurs very
early in mammals, where all of the meiotic and differentiation into oocytes before or just after birth, but ovulation does not take place until much later.
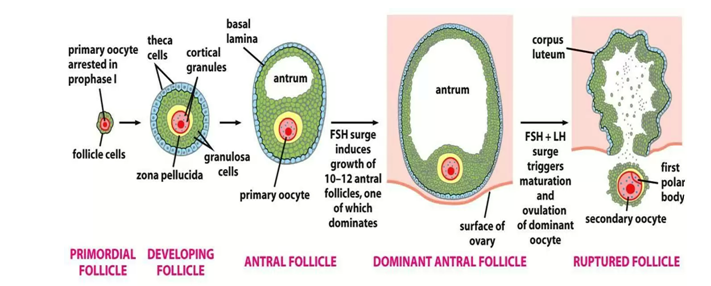
In any case the final production and delivery of the fully competent eggs or sperm require complex hormonal stimulation that occurs in adult, after the reproductive organs are fully mature.
If fertilization does not occur, corpus luteum regresses leaving a scarred area called corpus luteum albicans (white body). This leads to subsequent decrease in the level of progesterone.
As this happens, FSH is no longer inhibited and therefore its level increases in the blood. This marks the beginning of ovarian cycle
2. UTERINE CYCLE
Is a repeating series of changes in the structure of uterus.
It takes 21 – 35 days. It is divided into:-
Menses
Proliferation phase.
Secretory phase.
The Menstrual phase (menses)
This involves the shading of the epithelial lining of the endometrium. This phase and the process associated with it are explained as follows;
Following the regression of the corpus luteum, the level of progesterone in the blood decrease. This leads to the construction of spiral arteries that supply oxygen to the endometrium thus receive small amount of oxygen and consequently die.
By negative feedback, the spiral arteries then dilates allowing more blood to flow towards the dead cells in the uterine walls.
The pressure exerted by the blood causes the wall to disintegrate and flow out together with variable amount of blood in the menstrual flow. Usually endometrium sloughts out in patches.
Proliferation phase (Proliferative phase)
Involves the rapid proliferation multiplication of the endometrium under the influence of oestrogen from developing follicle.
Secretory phase
During this phase, progesterone from corpus luteum gland and this maintains the lining of the uterus in receptive state for implantation of the zygote.
Fig: Uterine cycle
DIFFERENCES
|
OESTRUS CYCLE
|
MENSTRUAL CYCLE
|
|
Common to lower mammals eg. Cats, dogs, caw etc.
|
– Is characteristics to the higher mammals of the order primate eg. Human beings, chimpanzee, gorilla, monkey etc.
|
|
In oestrus cycle, the endometrium cycle is absorbed if conception does not occur during the cycle.
|
-Animals with menstrual cycle shed the endometrium through menstruation.
|
|
In species with oestrus cycle, females are generally only sexually active during the oestrus cycle. This is referred to as in heat.
|
– Females of species with mentrual cycle can be sexually active at any time in their cycle even when they are not about to ovulate.
|
|
Period of heightened sexual activity conciding with ovulation is the most prominent event.
|
– Menstruation, the discharge of blood and uterine lining is the most prominent event.
|
|
Occurs less frequently eg. once per year.
|
– Occurs more frequently than oestrus cycle eg. once every month.
|
NOTE:–
Humans unlike some other species have once obvious external signs to signal receptivity at ovulation (concealed ovulation).
Research has shown however, that women tend to have more sexual thoughts and are most prone to sexual activity right before ovulation.
SIMILARITIES BETWEEN OESTRUS AND MENSTRUAL CYCLE
Both comprises of recurring physiological changes that are induced by reproductive hormones in most mammalian placental females.
Both start after puberty in sexually mature females and are interrupted by oestrus phases, continue until menopause.
METAMORPHOSIS
Definition:
Metamorphosis is the change of form of an organism in the course of its development.
Metamorphosis is caused by hormones from the brain and three other endocrine structures two of which are the corpus allatum and corpus cardiacum which are extensions of the brain.
BRAIN
This has neuro – secretory cells for secretion of the brain hormone (BH); This influences the secretion of ecdysone (a hormone controlling ecdysis) hormone from the pro-thoracic glands. The ecdysone hormone is stored in the thoracic gland.
CORPUS ALLATUM
This is an extension of the brain which secretes juvenile hormone (JH). This JH is dominant during the larval stage controls growth and moulting.
CORPUS CARDIACUM
It is also extension of the brain which basically receives brain hormone from the neuro-secretory cells of the brain, stores it before pouring it out.
PROTHORACIC GLAND
It secretes a prothoracic gland hormone (PGH) or ecdysone hormone. This is secreted when JH diminishes and it controls the process of population and emergency of an adult from the pupa. It is also controlling ecdysis.
TYPES OF METARMORPHOSIS
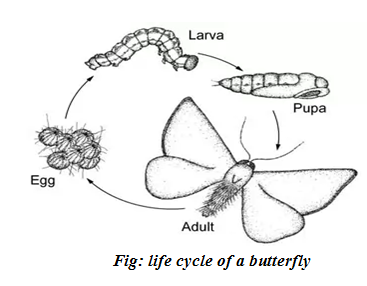
COMPLETE (holometabolous) METAMORPHOSIS
Type of metamorphosis in which four stages are involved
Example: – Housefly, butterfly.
Fig: life cycle of a butterfly
The embryo of an insect (for example a moth butterfly, beetle or fly) which undergo complete metamorphosis develops into a young form called a larva which appears very different from the adult.
Larvae lack many of the structures of the adult.
Butterfly larva has no wings and lacks compound eyes and jointed legs. They have become little more than feeding machines whose primary function is to find and consume food.
Once it has reached a certain size, the lava stops feeding and becomes a pupa by enclosing itself in the protean case within the case tissues are broken down and recognised so that they undergo a remarkable transformation to the adult form, the imago.
Once the adult has emerged from the case with fully developed wings, it can no longer moult. This restricts growth.
INCOMPLETE (hemimetabolous) METAMOPHORSIS
Is the type of metamorphosis in which an insect develops through three stages.
Example:- grasshopper, cockroach or locust undergo incomplete metamorphosis.
The embryo of an insect such as grasshopper, cocroach or locust undergo incomplete metamorphosis.
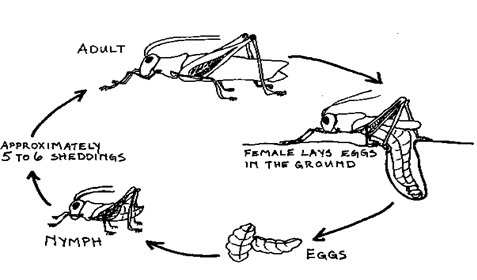
Fig: life cycle of a grasshopperIt develops into a nymph which closely resembles the adult form but which has a number of adaptive features which enables it to live in different habitat and eat different good from the adult.
In order to grow the nymph moult several times and go through a number of developmental stages called instars. The instar emerges as the adult with all the adult organs.
ADVANTAGES OF METAMORPHOSIS
Metamorphosis enables juvenile and adult forms to live different habitats and exploit different resources. This reduces competition between the different developmental stages.
Metamorphosis allows the larva and adult stages to become highly specialized for particular functions; usually the larval stage is specially adapted for feeding and the adult for reproduction.
REPRODUCTION IN FLOWERING PLANTS
– The reproductive structure of the flowering plant is the flower.
GAMETOGENESIS IN FLOWERING PLANTS
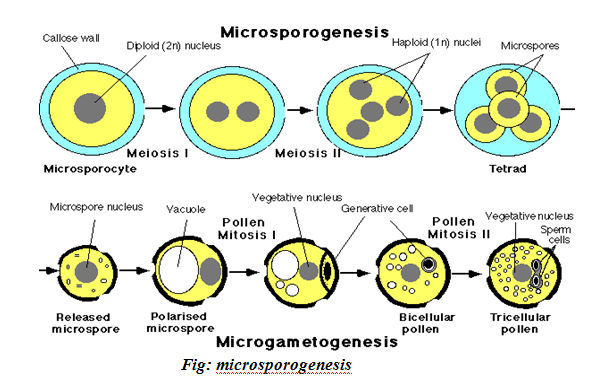
– as in animals, gametogenesis in flowering plants is the formation of gametes producing cells, the microspores s and the megaspores. The process whereby the microspores are produced is called Microsporogenesis where as megaspores are produced during megasporogenesis. The former forms pollen embryo female gamete.
DEVELOPMET OF POLLEN GRAINS: MICROSPOROGENESIS:
This occurs in the pollen sacs of the others, in these sacs each pollen or mother cell (2n) undergoes meiosis I to produce two haploid cells. Each of the resulting daughter cells undergoes meiosis II to produce a total of four haploid cells the four cells separate and each cell develops a thick wall over it. This wall is called an exine inside which lies an intine. Te pollen grain at this stage is equivalent to the microspore.
Its nucleus divides by mitosis to produce two nucleus the generation nucleus and the pollen tube nucleus
SEXUAL REPRODUCTION IN PLANT
Sexual reproduction in angiosperms occurs in the gametophyte generation. The structure for sexual reproduction is the flower.
It is within the flower where spore and gametes are developed.
GAMETOGENESIS
Occurs in two ways:-
(i) Microsporogenesis.
(ii)megasporogenesis.
MICROSPOROGENESIS
Is the process by which mature plant produces male gametes (pollen grains or microspores) at anthers of a flower.
The process takes place in the lobes of the anther.
MECHANISM:-
The microspore mother cell pollen mother cell 2n also called primary microsporocyte, undergo meiosis I to produce two haploid cells (dyad).
The products of meiosis I undergo meiosis II producing 4 cells (tetrad). The cells in the tetrad get separated from one another and cell (microspore pollen grain) secretes an additional wall over the present wall.
The nucleus of the pollen grain divides mitotically to produce two nuclei, the pollen tube, nucleus and the generative nucleus.
At this point the pollen grain and its contents if referred to as the male gametophyte because the male gametes will develop from the generated nucleus.
The mature pollen grain has two walls; the inner (INTINE) and the outer (EXINE). Exine has various pits (Sculptures).
MEGASPOROGENESIS
This is the development of the embryo sac (megaspore). The process takes place in the ovule of the ovary.
MECHANISM
The megaspore mother cell (2n) undergo meiosis to produce two cells (n) but only one continues to develop under the influence of nutrients from nucleus and become the embryo sac.
The nucleus of the embryo sac divides mitotically three times to produce 8 nuclei the antipodals migrate, polar nuclei remain at the centre and 3 nuclei the female gamete and sinergid migrate to the microphylen end.
Soon after mitosis the embryo sac and its content is referred to as the female gametophyte because one of the nuclei is the female gamete.
Fig: Development of embryo sac and female gamete.
Fig: Is carpel at fertilization. Note that, ovule which becomes the seed after fertilization, contains both diploid parent tissue and haploid embryo sac tissue.
DOUBLE FERTILIZATION AND ITS CONSEQUENCES
Double fertilization is a unique characteristic of the angiosperm. It is a result of multinucleate of the pollen grain and embryo sac.
Definition: Double fertilization is a type of fertilization which occurs in the flowering plant where the two types of nuclear fusion take place.
First: The male gamete nucleus fuses with the female gametes of the embryo sac to form the zygote (2n).
Second: The second male gamete fuses with diploid nucleus (resulting from the fusion of 2 polar nuclei) forming a primary endosperm (3n).
MECHANISM OF DOUBLE FERTILIZATION
The process of double fertilization is proceeded by pollination during which the pollen grain lands on the stigma.
As the pollen grain lands on the stigma the style tissue stars to secrete sugary solution including sucrose solution. The solutions are absorbed by the pollen grain which consequently swells.
As a result of swelling; the intine wall grows through the exile via one of the pits as pollen tube under the control of pollen tube down the style.
As the pollen tube continues to grow, the generative nucleus in the pollen grain divides by mitosis to produce two male gametes. In embryo sac, the two pollen nuclei fuse producing a diploid nucleus and the antipodal as the sinergid degenerate.
The embryo sac thus remains with only two nuclei an ovum (n) and a diploid nucleus resulting from the fusion of the polar nuclei.
The pollen tube continues to grow chemotactically towards the embryo sac under influence of chemicals secreted by the embryo sac. As the pollen tube reaches the micropyle, the following occurs in the pollen tube:-
The tip of the pollen tube bursts.
The contents of the pollen grain are discharged into the vicinity of the embryo sac.
Fertilization occurs following the discharge of contents of pollen grains. It is a double fertilization
How?
lower-roman;text-decoration: none;vertical-align: baseline”>
The haploid male gamete fuses with the female gamete to form a diploid zygote nucleus.
lower-roman;text-decoration: none;vertical-align: baseline”>
The second haploid male gamete fuses with the diploid (resulting) from the fusion of two polar nuclei to form a triploid primary endosperm nucleus.
TOPIC 3 : REPRODUCTION (I) ~ BIOLOGY FORM 6
TOPIC 4: GENETICS ~ BIOLOGY FORM 6


























")




 March 2024 – Primary School Exam")

 MARCH 2024")
, March 2024")
, March 2024")



 March 2024")
 – All Subjects")
 Joining Instruction & Courses Offered | Apply Now")




 SIGNIFICANCE OF MEIOSIS:
SIGNIFICANCE OF MEIOSIS:








")





 And Advanced Level (Form 5 and 6) All Subjects")

 TIMETABLE, MAY 2024")



 PDF")








Hi, There’s no doubt that your blog might be having internet browser compatibility issues. When I look at your web site in Safari, it looks fine however, when opening in IE, it’s got some overlapping issues. I simply wanted to give you a quick heads up! Other than that, excellent website!
Hi there! This post couldn’t be written any better! Going through this post reminds me of my previous roommate! He always kept preaching about this. I’ll forward this article to him. Fairly certain he’s going to have a good read. Thanks for sharing!
I’m amazed, I have to admit. Seldom do I encounter a blog that’s both educative and amusing, and without a doubt, you’ve hit the nail on the head. The problem is something that not enough men and women are speaking intelligently about. I am very happy I stumbled across this during my hunt for something concerning this.